Dynamique des populations
3. LA MORTALITÉ
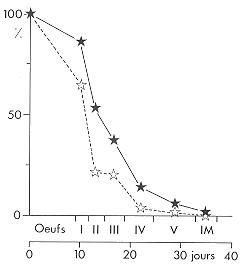 Courbes de survie observées dans la nature pour les stades embryonnaire et larvaire du Criquet migrateur Locusta migratoria capito, dans le Sud-Ouest de Madagascar (d'après M. LECOQ, 1975). Courbes de survie observées dans la nature pour les stades embryonnaire et larvaire du Criquet migrateur Locusta migratoria capito, dans le Sud-Ouest de Madagascar (d'après M. LECOQ, 1975).
En abscisse sont indiqués les stades embryonnaires (œuf), larvaire (de I à V) et imaginal (à téguments mous IM) ainsi que la durée du développement en jours.
En ordonnée figure le pourcentage d'individus survivants calculé à la moitié de chaque stade.
Les deux courbes concernent des résultats obtenus sur deux stations différentes. Le nombre de jeunes imagos survivant en fin de développement embryonnaire et larvaire ne représente qu'une faible fraction (1 à 2 %) de la population embryonnaire initiale.
La mortalité est un facteur de décroissance de la population. Elle survient à tout âge : état embryonnaire, état larvaire, état imaginal. Elle peut être naturelle (vieillissement) ou accidentelle. Dans ce dernier cas, la mort résulte des effets de facteurs abiotiques (inondation, sécheresse) ou de facteurs biotiques (prédateurs, parasites, maladies).
Les facteurs de mortalité peuvent être :
intrinsèques : ils sont liés à l'aptitude à survivre d'un individu, d'une population ou d'une espèce dans un milieu proche de son optimum écologique.
extrinsèques : ils dépendent de la résistance des organismes aux agressions extérieures : météorologie, parasitisme, prédatisme, maladies, compétition.
Leurs effets diffèrent considérablement selon les espèces, les acridiens à faible fécondité ayant généralement une meilleure aptitude à survivre dans leur milieu habituel que celles à fécondité très élevée.
Une distinction doit être faite entre la longévité physiologique et la longévité écologique. La
longévité physiologique rend compte de l'aptitude intrinsèque d'une espèce à vivre dans un milieu suboptimal. On l'oppose à la notion de longévité écologique qui est la durée réelle de vie observée chez une population naturelle.
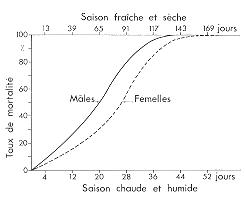 |
Courbe de mortalité imaginale du Criquet migrateur malgache Locusta migratoria capito, dans la nature (modifié d'après M. LAUNOIS & M.H. LAUNOIS-LUONG, 1980).
L'aspect genéral de la courbe de mortalité reste le même en saison chaude et humide comme en saison fraiche et sèche mais la longévité est plus grande au cours de cette dernière période de l'annee (78 jours en moyenne pour 50 % de la population, contre 24 jours).
|
Le taux de mortalité (Tm) est calculé par le rapport du nombre de morts (M) au nombre d'individus de la population entière (P), multiplié par 100 pour obtenir une valeur en pourcentage. Il a une expression comparable dans son principe à celle du taux de natalité.
Les tables de vie sont conçues pour suivre l'évolution du taux de mortalité d'une population. On estime à intervalles réguliers le nombre d'individus survivants du stade œuf au stade ailé. Il est également possible de construire une courbe de vie pour représenter l'évolution temporelle du pourcentage de criquets survivants en fonction de leur âge. Chez Schistocerca gregaria, la mortalité la plus importante se situe pendant les deux premiers stades larvaires. Elle s'atténue ensuite pour atteindre un minimum chez les jeunes imagos.
Les causes de mortalité sont différentes selon qu'il s'agit d'œufs, de larves ou d'ailés.
Tous les œufs pondus par une femelle ne sont généralement pas viables mais le pourcentage d'œufs qui ne se développent pas est souvent faible (quelques %) et relativement constant. Les œufs d'acridiens déposés dans le sol peuvent être détruits par de nombreux parasites et
prédateurs.
Les œufs sont également soumis aux aléas de leur environnement physico-chimique. Ils ont besoin d'eau pour se développer, qu'il y ait ou non induction de diapause. Une dessication ou une inondation survenant pendant un temps trop long provoque la mort par dessication ou asphyxie. L'approvisionnement en eau du sol doit donc être régulier et suffisant.
La mortalité post-embryonnaire survient après l'éclosion. Elle a lieu principalement pendant les trois premiers stades larvaires. L'une des causes majeures est le prédatisme. La prédation lors des mues n'est pas négligeable, en raison de la faiblesse des acridiens à ce moment-la.
Les parasites et les prédateurs des larves et des ailés ne sont pas généralement les mêmes que ceux des œufs. La distribution des ennemis naturels, ceux qui ménagent à court terme la vie de l'hôte en vivant à ses dépens (parasite) ou le tuent en le consommant (prédateur), est rarement homogène.
Les acridiens sont aussi victimes de maladies. Des épidémies foudroyantes et dévastatrices (épizooties) peuvent détruire localement des populations entières. Les agents pathogènes sont des micro-organismes appartenant à trois groupes principaux : les protozoaires, les champignons et les bactéries.
 Influence de la pluviométrie du mois d'observation et du mois précédent sur le taux de développement des populations imaginales du Criquet migrateur Locusta migratoria capito, dans le Sud-Ouest de Madagascar (d'après M. LAUNOIS, 1974e). Influence de la pluviométrie du mois d'observation et du mois précédent sur le taux de développement des populations imaginales du Criquet migrateur Locusta migratoria capito, dans le Sud-Ouest de Madagascar (d'après M. LAUNOIS, 1974e).
Le taux de développement des populations d'ailés est exprimé en pourcentage. Ce graphique révèle l'importance prépondérante de la pluviométrie du mois d'observation sur celle du mois précédent. Le niveau des populations imaginales est donc plus dépendant des conditions actuelles d'environnement, que des
conditions passées. Cela tient à la rapidité de développement de cette espèce et à ses capacités d'ajustement au
milieu par deplacement adaptatif. Son optimum de développement est le plus souvent réalise dans des régions
soumises à une pluviométrie comprise entre 50 et 100 mm d'eau par mois.
Les variations de mortalité tiennent surtout aux changements brusques des conditions d'environnement des espèces et aux possibilités d'ajustement ou d'adaptation de ces dernières. La survie des stades les plus vulnérables dépend des conditions locales. On admet généralement que la moitié des individus disparaissent à chaque stade.
|

