Morphologie
2. LE TÉGUMENT
Le tégument est le revêtement externe du corps. Il est généralement très dur et agencé en
segments articulés les uns aux autres. Il comprend une couche de cellules hypodermiques et
des produits de sécrétions formant la cuticule.
 |
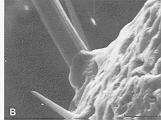
Reconstitution schématique du tégument en trois dimensions.
(d'après A.G. RICHARDS, 1951).
c : cuticule,
ca : canal glandulaire,
cg : cellule glandulaire,
cto : cellule tormogène,
ctr : cellule trichogène,
e : épiderme,
en : endocuticule,
ép : épicuticule,
ex : exocuticule,
m : membrane basale,
p : procuticule,
s : soie.
|
2.1. STRUCTURE, COMPOSITION ET FONCTIONS DU TÉGUMENT
La cuticule forme un véritable squelette externe articulé grâce à l'existence de zones souples entre les différentes pièces sclérifiées. Des invaginations au niveau de certaines sutures forment des apodèmes sur lesquels s'insèrent des muscles. Cette carapace impose un mode de croissance particulier par mues successives.
La cuticule est formée de différentes couches superposées qui sont, de l'extérieur vers l'intérieur du corps :
l'épicuticule, couche très fine (1 micron) comprenant essentiellement des cires,
l'exocuticule,
l'endocuticule, qui avec l'exocuticule forme la procuticule.
Ces strates sont déposées par les cellules de l'épiderme. La membrane basale, produite par ces mêmes cellules du côté interne du corps, les isole de la cavité générale où se trouvent les organes vitaux.
Le tégument présente souvent des ornementations telles que des piquants, des épines, des spicules qui sont des phanères fixes. On les oppose souvent aux phanères mobiles (soies sensorielles, soies glandulaires). Les phanères fixes sont souvent de simples épaississements de la cuticule. Les phanères mobiles sont de structure plus complexe. Des cellules épidermiques spécialisées, sensorielles ou sécrétrices, entrent dans leur formation.
|
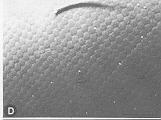
Coupe transversale du tégument du Criquet pèlerin, Schistocerca gregaria, au niveau des tergites abdominaux d'une larve de dernier stade
(d'après S.R.A. MALEK, 1958).
e : épiderme,
ee
: espace exuvial,
em : partie mélanisée de l'endocuticule,
en : endocuticule,
ep : épicuticule,
ex : exocuticule,
m : membrane basale,
me : membrane ecdysiale,
o : nocyte,
pe :endocuticule pré-exuviale.
|

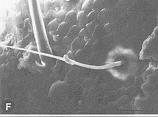
|
| A : |
larve venant de muer |
| B : |
2 heures après la mue |
} dépôt des couches de l'endocuticule et des stries d'accroissement |
| C : |
3 à 4 heures |
| D : |
4 à 5 jours, formation de l'espace exuvial |
| E : |
5 à 6 jours |
} préparation de la nouvelle cuticule |
| F : |
6 à 7 jours |
| G : |
peu de temps avant la mue imaginale |
La préparation de la mue commence plusieurs jours avant le jour d'exuviation par la formation d'une nouvelle cuticule sous l'ancienne (A, B, C). Vers le milieu du stade D, l'ancienne cuticule se sépare de l'épiderme et le jour suivant, une nouvelle cuticule apparaît sur la face externe de l'épiderme. Sur la face interne de l'ancienne cuticule, se forme la membrane ecdysiale. L'espace compris entre la nouvelle et l'ancienne cuticule (espace exuvial) s'aggrandit. L'ancienne endocuticule est digérés par les sécrétions des glandes de mue situées dans
l'épiderme. La membrame ecdysiale persiste. Elle est rejetée lors de la mue avec le reste de l'ancienne cuticule.
La procuticule constitue la partie la plus importante du tégument. Elle est chargée de protéines et de chitine. La chitine, polysaccharide aminé de poids moléculaire élevé, assez
proche de la cellulose, est absente de l'épicuticule. La dureté de la cuticule provient du tannage
de la fraction protéique.
Chaque couche cuticulaire a un rôle particulier :
l'épicuticule assure l'imperméabilité,
l'exocuticule la rigidité des parties dures,
l'endocuticule la flexibilité et la solidité du tégument.
Le tégument comprend des sclérites (parties dures) et des membranes souples. Les sclérites portent le nom de tergites sur le dos, de pleurites sur les côtés, de sternites sur le ventre. Sur
le thorax, les ailes s'insèrent entre les tergites et les pleurites ; les pattes entre les pleurites et les sternites. L'abdomen ne possède pas de pleurite.
L'ensemble du tégument a plusieurs fonctions :
une protection mécanique du corps qu'il enrobe étroitement,
une barrière physico-chimique vis-à-vis de nombreuses substances toxiques mais aussi dans les échanges gazeux,
un rôle d'accumulation et de conservation de la chaleur,
une fonction d'organe de réserves utilisables en cas de diète prolongée,
enfin, les différenciations internes et externes du squelette permettent aux organes d'être maintenus en place et à l'insecte de se mouvoir grâce à l'insertion des muscles sur des parties dures.
2.2. LA PIGMENTATION DU TÉGUMENT
La coloration du tégument provient de deux sources fondamentalement différentes :
une source pigmentaire, c'est-à-dire que la teinte observée est d'ordre physique sur support chimique,
l'autre est physique. Elle correspond à une réponse sélective de certaines parties de la cuticule à la lumière incidente.
Les pigments les plus communs sont:
les ommochromes : granules bruns, jaunes, rouges, violets,
les caroténoïdes : granules jaunes et rouges,
les ptérines : granules blancs, rouges, jaunes, verts.
Les pigments cuticulaires sont essentiellement formés de mélanine avec des granules jaunes, rouges, bruns et noirs. Ces pigments sont le plus souvent des protéines spécifiques qui peuvent parfois être d'origine végétale (chlorophylles, caroténoïdes, anthocyanes) et sont alors stockés après ingestion des plantes tout en restant intacts.
Les teintes les plus communes sont le vert, le gris et les bruns, avec des extrêmes jaune-paille et noir. Les couleurs vives, rouge, bleu et jaune, sont souvent limitées aux ailes et aux fémurs postérieurs. Elles sont plus rares sur les autres parties du corps.
La pigmentation change en fonction de facteurs dont l'action diffère selon les espèces :
facteurs internes : état de maturité sexuelle, densité de population,
facteurs externes : température, humidité, taux de gaz carbonique, photopériode, couleur de l'environnement.
Les modifications de coloration surviennent lors des mues lorsque les cellules épidermiques reprennent leur activité ou par migration de pigments intracellulaires. Il existe des formes mutantes comme les albinos, suite à une abscence de pigmentation.
Les larves et les adultes d'une même espèce peuvent présenter plusieurs types pigmentaires :
Pnorisa carinata, avec ou sans bandes latérales brunes sur le pronotum,
Acridoderes strenuus, avec écusson pronotal apparent ou non,
chez de nombreux Oedipodinae, polychromisme vert/brun souvent rencontré.
Certaines espèces peuvent montrer des différences pigmentaires en fonction des taxons géographiques : Acrotylus patruelis à Madagascar a deux couleurs d'ailes postérieures (jaune ou rouge), alors qu'en Afrique, la forme jaune est exceptionnelle.
Les conditions de développement influent souvent sur les types pigmentaires. Ainsi, la couleur verte de Locusta migratoria est-elle fréquente en saison des pluies, alors que la couleur brune
domine en saison sèche.
La pigmentation générale peut changer au cours de la vie : premiers stades larvaires bruns, puis derniers stades verts. L'amplitude des variations de coloration dépend de l'état phasaire des espèces grégariaptes. Les criquets en phase solitaire présentent des colorations variées, ajustées à leur environnement du moment, bien davantage que les criquets en phase grégaire.
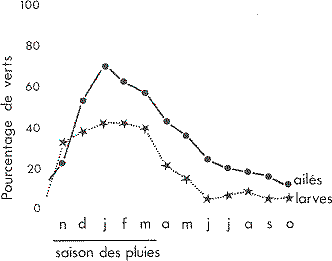
Évolution du taux de larves et d'ailés de coloration génerale verte du Criquet migrateur Locusta migratoria dans les clairières du Plateau Mahafaly du
Sud-Ouest de Madagascar.
Moyenne sur 8 ans 1965-1973. Extrait des archives du
Service antiacridien de Madagascar).
À aucun moment de l'année, tous les individus d'une population ne sont verts ou bruns. Le pourcentage de criquets verts est plus important en saison des pluies qu'en saison sèche il croît plus rapidement qu'il ne diminue. Les larves réagissent les premieres en début de saison des pluies, mais la proportion d'individus verts est généralement plus faible que chez les ailés. Les larves de premier stade ont été exclues du calcul car elles sont toujours brunes chez cette espèce.
Sur le plan de l'identification des espèces, l'usage des types pigmentaires est relativement limité car ils sont sujets à de trop grandes variations.
|