Physiologie
5. LA FONCTION DE REPRODUCTION
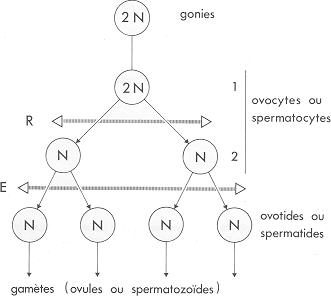 Schéma général de la méïose Schéma général de la méïose
La reproduction sexuée est fondée sur deux phénomènes complémentaires : la réduction chromatique (ou méïose) et la fécondation.
La méïose comporte deux divisions successives : une mitose réductionnelle (R) et une mitose équationnelle (E). Elle conduit à la transformation des gonies (ou cellules indifférenciées) en gamètes qui sont des cellules sexuelles à garniture chromosomique simple (cellule haploïde à n chromosomes). Les gamètes femelles (ou ovules) sont généralement peu mobiles et pourvus de réserves abondantes. À l'inverse, les gamètes mâles (ou spermatozoïdes) sont très mobiles et presque dépourvus de réserves.
La fécondation résulte de la fusion d'un ovule et d'un spermatozoïde, il se forme alors un œuf (ou zygote), cellule munie d'une double garniture chromosomique (cellule diploide à 2n chromosomes). L'œuf se divise ensuite par mitose et la différenciation cellulaire permet de former un nouvel organisme complet.
Le développement d'un œuf qui n'a pas été fécondé relève de la parthénogenèse. Chez les acridiens, la duplication des chromosomes des œufs vierges des femelles, par suite du déterminisme génétique du sexe, conduit à une descendance femelle : la parthénogenèse est dite thélythoque. La majorité des acridiens ne pratique pas habituellement ce mode de reproduction sans male.
La reproduction de type sexué repose sur deux phénomènes biologiques fondamentaux :
la méïose ou division cellulaire avec réduction chromatique (division par 2 du nombre de chromosomes caractéristiques de l'espèce) amenant la production de cellules reproductrices mâles et femelles à n chromosomes,
la fécondation conduisant à la fusion de deux cellules reproductrices mâle et femelle pour former un œuf ou zygote à 2n chromosomes, qui se développe ultérieurement en organisme par multiplication (mitose) et différenciation cellulaires.
L'intérêt de la reproduction sexuée réside dans la possibilité de recombiner les caractères parentaux.
La structure des tubes séminifères est bien connue ainsi que la formation des spermatozoïdes. On sait que les sécrétions qui accompagnent les gamètes mâles dans le spermatophore ont un rôle activateur important du métabolisme des femelles inséminées. Par contre, les fonctions du complexe phallique sont mal comprises. L'érection du pénis est produite par un apport local d'hémolymphe et le jeu de muscles viscéraux particuliers.
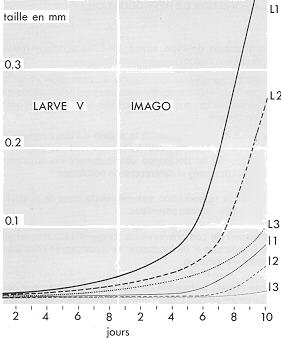 |
Évolution comparée de la taille des 3 premiers ovocytes des ovarioles de Locusta migratoria cinerascens (souche Hyères) au laboratoire depuis le premier
jour de la larve de 5e stade jusqu'au 11e jour après la mue imaginale (d'après M. LAUNOIS, 1968, non publié)
Les deux dimensions mesurées, la longueur et la largeur des ovocytes, montrent que la taille de chacun des 3 premiers ovocytes est légèrement différente dès avant la mue imaginale. Un écart plus important apparaît ensuite, d'abord progressivement avec le dépôt d'un vitellus blanc, puis plus rapidement apres le 5e jour avec le dépot du vitellus jaune en premier lieu dans le 1er, puis dans le 2e et enfin dans le 3e ovocyte. Le nombre de jours séparant la croissance de ces 3 ovocytes est sensiblement égal au
rythme de ponte dans les conditions d'observation.
L1, L2, L3 : longueur des ovocytes,
I1, I2, I3 : largeur des ovocytes.
|
Le fonctionnement des ovaires a fait l'objet d'un grand nombre de recherches, à cause de la nécessité de connaître la production des œufs pour estimer le potentiel d'expansion d'une espèce.
Les ovaires sont composés d'ovarioles. Le nombre total de ces derniers est fixé dès la naissance. Chaque ovariole est une unité structurale fonctionnelle. L'ovocyte situé le plus près du point d'insertion sur le calice est le premier qui subit le dépôt des réserves vitellines jaunâtres. L'ovocyte sus-jacent n'entreprend sa croissance que lorsque le précédent a presque achevé la vitellogenèse, et ainsi de suite jusqu'au cinquième ovocyte au moins où une différence de taille selon le rang est encore visible.
Les ovocytes subissent une croissance vitelline en deux étapes :
dépôts blancs, d'aspect laiteux,
dépôts jaunes.
lls peuvent parvenir à maturation ou subir un arrêt de croissance à tout moment de la vitellogenèse et jusqu'à la formation du chorion. Chaque ovocyte est entouré d'un manchon de cellules folliculaires. Ces cellules changent d'activité en fin de vitellogenèse en sécrétant l'enveloppe externe du futur œuf, le chorion.
En cas d'arrêt de la vitellogenèse lorsque les ressources en nutriments sont limitées ou le métabolisme maternel perturbé, l'ovocyte subit une résorption vitelline. Celle-ci se traduit par une diminution de taille due à la libération dans l'hémolymphe de métabolites réutilisables, et par une pigmentation plus vive rouge-orangée, par suite de la concentration de produits caroténoïdes.
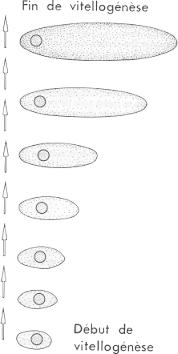 Évolution de la position du noyau au sein de l'ovocyte en fonction de la croissance de ce dernier chez le Criquet migrateur Locusta migratoria. Évolution de la position du noyau au sein de l'ovocyte en fonction de la croissance de ce dernier chez le Criquet migrateur Locusta migratoria.
Pendant la période de prévitellogenèse, le noyau est centré dans l'ovocyte. Lorsque ce dernier s'allonge et que les cellules folliculaires se multiplient, la vitellogenèse est amorcée. Le noyau va alors se déplacer progressivement vers le futur pôle animal de l'ovocyte qui acquiert ainsi sa polarité.
Lorsque l'ovocyte a achevé sa croissance avec la formation du chorion, il est expulsé dans les calices dans l'attente d'une ponte imminente. C'est l'ovulation.
Si l'ovocyte échoue dans sa croissance, le corps de résorption est formé d'une enveloppe folliculaire avec des déchets vitellins colorés en orangé ou en rouge.
L'aspect général des ovarioles diffère chez les jeunes femelles et chez celles qui ont pondu de nombreuses fois. Chez ces dernières, la gaine conjonctive est distendue et les ovocytes en prévitellogenèse sont souvent plus gros que ceux observés chez les jeunes reproductrices.
L'activité des ovaires est sous contrôle hormonal.
L'état physiologique des parents influe sur la viabilité des œufs et, spécifiquement chez les locustes, sur la transmission de certains caractères phasaires à la descendance.
|

